INTRODUCTION
Improved animal health and performance always has been the goal of people associated with livestock production. Consequently, any feedstuff, feed additive, drug or other compound that is capable of enhancing animal health or performance will interest producers, veterinarians, and animal nutritionists. Several compounds have been used to improve animal performance either by manipulation of the rumen environment (e.g., sodium bicarbonate) or by directly altering the composition and metabolic activities of rumen microorganisms (e.g., ionophores).
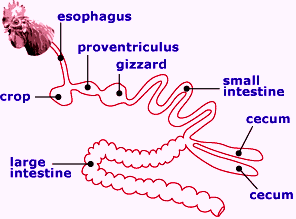
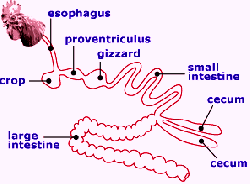
Digestion is the process of breaking down large, complex molecules, as provided by the birds’ feed, into smaller components that can be absorbed into the portal blood system. The process involves changes in both physical and chemical structures of most dietary components. Poultry feeds consist of a complex array of particles differing not only in chemical composition, but also in size, hardness, solubility and ionic characteristics.
Under ideal conditions, this array of particles and chemicals with different characteristics degrade slowly in a step-wise manner as feed passes from the mouth to the large intestine. Particle breakdown is a constant process, although the gizzard provides the major site of this activity.
Enzymes are largely responsible for molecular degradation, although their pH greatly influences their efficacy.
When digestion is reduced, there will be reduced bird growth and/or increased feed intake. Indigestion may also cause problems with manure/litter management, because non-digested residues in the large intestine often adsorb more water or produce feces that are more viscous.
Animal feed contains Cereals and Cakes.
Cell walls of cereals are primarily composed of carbohydrate complexes referred to as Non Starch Polysaccharides (NSP).
ANFs present in these NSP (like ß-glucans and arabinoxylans) are non digestible and form high-molecular-weight viscous aggregates in the gastrointestinal tract.
They
Affect the digestive enzymes.
Cause endogenous losses
Reduce the rate of passage.
Stimulates pathogenic microbial proliferation.
Enzymes such as xylanase or ß-glucanase into diets having ANF can effectively decrease viscosity and consequently reduce the anti nutritional effect of NSP.
Cellulase, ß-Glucanase, Xylanase, and Pectinase can degrade plant origin cell wall polymers.
Amylase can increase the gut absorption levels of starches.
Hemi Cellulase can degrade the difficult fiber.
Protease can degrade the proteins.
Lipase can degrade the lipids.
Phytase helps in solubilising the phyto phosphorous.
Tannases can degrade the plant toxins.
|
G.I Tract Region |
Enzyme (or secretion) |
Substrate |
End Product |
pH |
|
Mouth |
Saliva |
Lubricates and softens food |
|
|
|
Crop |
Mucus |
Lubricates and softens food |
4.5 |
|
|
Stomach (Gizzard and proventiculus) |
HCI |
Lowers stomach pH |
2.5 |
|
|
Pepsin |
Protein |
Polypeptides |
||
|
Duodenum |
Trypsin, Chymotrypsin and Elastases |
Proteins, Peptones and Peptides |
Peptones, Peptides and amino acids |
|
|
Carboxy-petidases |
Peptides |
Peptides and amino acids |
6 |
|
|
Collagenase |
Collagen |
Peptides |
to |
|
|
|
|
|
6.8 |
|
|
Jejunum |
Peptidases |
Peptides |
Dipeptides and amino acids |
5.8 to 6.6 |
Posttranslational glycosylation has been reported to protect enzymes from deactivation caused by high temperatures and proteinases
(Olsen and Thomsen, 1991).
All enzyme feed additives are considered either food additives or GRAS substances
The activity of enzymes retains more than 95% when stored at the temperature of 25 Deg C upto 3 months
PROTEINS:
Protein and amino acid availability are of greatest concern in animal and vegetable protein ingredients. Protein content and availability from cereals and their by-products seem to be more consistent and little affected by processing conditions.
|
Feedstuff |
C. Protein (%) |
Digestibility (%) |
|||
|
|
|
C. Protein |
Lys |
Met |
Cys |
|
Vegetable sources (cereals) |
|||||
|
Yellow maize |
8 |
82 - 86 |
81 |
91 |
85 |
|
Wheat |
12 |
78 - 82 |
81 |
87 |
87 |
|
Barley |
10 |
70 -82 |
78 |
79 |
81 |
|
Sorghum |
10 |
67 - 72 |
78 |
89 |
83 |
|
Vegetable sources (oil seed meals) |
|||||
|
Peanut meals |
49 |
88 - 91 |
83 |
88 |
78 |
|
Soybean meals |
46 |
83 - 87 |
91 |
92 |
82 |
|
Cottonseed meal |
43 |
61 - 76 |
67 |
73 |
73 |
|
Animal sources |
|||||
|
Blood meal |
88 |
82-92 |
86 |
91 |
76 |
|
Fish meal |
66 |
86 - 90 |
88 |
92 |
73 |
|
Meat meal |
60 |
75 - 80 |
79 |
85 |
58 |
|
Feather meal |
87 |
36 - 77 |
66 |
76 |
59 |
FATS:
|
G.I Tract Region |
Enzyme (or secretion) |
Substrate |
End Product |
pH |
|
Mouth |
Saliva |
Lubricates and softens food |
|
|
|
Crop |
Mucus |
Lubricates and softens food |
4.5 |
|
|
Stomach (Gizzard and proventiculus) |
HCI |
Lowers stomach pH |
2.5 |
|
|
|
||||
|
Lipase |
Fats |
Fatty acids, mono-glycerides and glycerol |
||
|
Duodenum and Jejunum |
Bile |
Fats |
Emulsification |
|
|
Lipase |
Fats |
Fatty acids, mono-glycerides and glycerol |
5.8 |
|
|
Cholesterol esterase |
Fatty acid - cholesterol esters |
Fatty acid, cholesterol |
to |
|
|
|
|
|
6.6 |
|
|
|
Digestibility |
Metabolizable energy (kcal/kg) |
||
|
Fat Type/Age |
0-21d |
>21d |
0-21d |
>21d |
|
Tallow |
80 |
86 |
7400 |
8000 |
|
Poultry Fat |
88 |
97 |
8200 |
9000 |
|
Fish oil |
92 |
97 |
8600 |
9000 |
|
Vegetable oil |
95 |
99 |
8800 |
9200 |
|
Coconut oil |
70 |
84 |
6500 |
7800 |
|
Palm oil |
77 |
86 |
7200 |
8000 |
|
Vegetable soapstock |
84 |
87 |
7800 |
8100 |
|
Restaurant grease |
87 |
96 |
8100 |
8900 |
Cellulase
Breaks down cellulose and chitin
(chitin is cellulose like fiber found in the cell wall of candida).
Cellulases acts on cellulose molecules by hydrolysing the beta-1, 4 glycosidic linkages.
They largely produces cellobiose, which can ultimately yield glucose units, depending on
the characteristic of the enzyme.
It helps free nutrients in both fruits and vegetables.
Measured in CU (Cellulase Units).
Activity of Enzyme:
One activity u/mg (u/ml) of cellulase is defined as that quantity of enzyme that can liberate 1g glucose in one minute at pH of 4.8.
Standard Product: 8000 u/g
Use the preparation between pH of 4.0~5.0,
Generally speaking, adding the preparation 50g to 1t dry materials will play well.
Protease:
Bonds with alpha 2-macroglobulin to support immune function when taken on an empty stomach.
Protease is responsible for digesting proteins in food, which is probably one of the most difficult substances to metabolize. Because of this, protease is considered to be one of the most important enzymes that we have. If the digestive process is incomplete, undigested protein can wind up in the circulatory system, as well as in other parts of the body.
When protease is present in higher quantities, it can help to clean up the body by removing the unwanted protein from the circulatory system. This will help to clean up the blood stream, and restore the energy and balance.
One of the tricks of an invading organism is to wrap itself in a large protein shell that the body would view as being "normal". Large amounts of protease can help remove this protein shell, and allow the body`s defense mechanisms to go into action. With the protective barrier down, the immune system can step in and destroy the invading organism.
Protease refers to a group of enzymes whose catalytic function is to hydrolyze (breakdown) peptide bonds of proteins. They are also called proteolytic enzymes or proteinases. Proteases differ in their ability to hydrolyze various peptide bonds. Each type of protease has a specific kind of peptide bonds it breaks. Examples of proteases include: fungal protease, pepsin, trypsin, chymotrypsin, papain, bromelain, and subtilisin.
Proteolytic enzymes are very important in digestion as they breakdown the protein foods to liberate the amino acids needed by the body. Additionally, proteolytic enzymes have been used for a long time in various forms of therapy. Their use in medicine is gaining more and more attention as several clinical studies are indicating their benefits in oncology, inflammatory conditions and immune regulation.
Contrary to old beliefs, several studies have shown that orally ingested enzymes can bypass the conditions of the GI tract and be absorbed into the blood stream while still maintaining their enzymatic activity. Commercially, proteases are produced in highly controlled aseptic conditions for food supplementation and systemic enzyme therapy. The organisms most often used are Aspergillus niger and oryzae.
Measured in HUT (Hemoglobin Units in a Tyrosine Base).
Lipase
Lipase is an enzyme necessary for the absorption and digestion of nutrients in the intestines. This digestive enzyme is responsible for breaking down lipids (fats), in particular triglycerides, which are fatty substances in the body that come from fat in the diet. Once broken down into smaller components, triglycerides are more easily absorbed in the intestines. Lipase is primarily produced in the pancreas but is also produced in the mouth and stomach. Most people produce sufficient amounts of pancreatic lipase.
Along with lipase, the pancreas secretes insulin and glucagon, hormones that the body needs to break down sugar in the bloodstream. Other pancreatic enzymes include amylase, which breaks down amylose (a form of starch) into its sugar building blocks, and protease, which breaks down protein into single amino acids.
Source:
Lipase, monoacylglycerol:
Penicillium camembertii
Lipase, triacylglycerol:
Aspergillus niger
Aspergillus oryzae
Aspergillus oryzae, containing the gene for Lipase, triacylglycerol isolated from Fusarium oxysporum
Aspergillus oryzae, containing the gene for Lipase, triacylglycerol isolated from Humicola lanuginosa
Aspergillus oryzae, containing the gene for Lipase, triacylglycerol isolated from Rhizomucor miehei
Rhizopus arrhizus
Rhizomucor miehei
Rhizophus niveus
Rhizophus oryzae
Uses
In general, lipase supplements are thought to help the body absorb food more easily, keeping nutrients at appropriate, healthy levels throughout the body. Studies suggest that they may also be helpful for the following conditions:
Celiac Disease
Pancreatic enzymes have been most studied as part of the treatment for celiac disease. Celiac disease is a condition in which dietary gluten causes damage to the intestinal tract. Symptoms include abdominal pain, weight loss, and fatigue. People with celiac disease must consume a life-long gluten-free diet. Lipase, along with other pancreatic enzymes, may help in the treatment of this condition by enhancing the benefit of a gluten-free diet. In a study of 40 children with celiac disease, for example, those who received pancreatic enzyme therapy (including lipase) demonstrated a modest increase in weight compared to those who received placebo. The improvement in weight occurred within the first month of use; taking the pancreatic enzyme supplements for an additional month did not lead to more weight gain.
Indigestion
In a small study including 18 subjects, supplements containing lipase and other pancreatic enzymes were found to reduce bloating, gas, and fullness following a high-fat meal. Given that these symptoms are commonly associated with irritable bowel syndrome, some with this condition may experience improvement with use of pancreatic enzymes.
Other
Although scientific evidence is lacking, lipase has been used by trained clinicians to treat food allergies, cystic fibroris, and autoimmune disorders, such as rheumatoid arthritis and lupus.
Dietary Sources
Lipase is produced primarily in the pancreas and is not found in food.
Available Forms
Lipase supplements are usually derived from animal enzymes, although plant sources of lipase and other digestive enzymes have become increasingly popular. Lipase may be taken in combination with protease and amylase enzymes.
Pectinase
Breaks down carbohydrates, such as pectin found in many fruits and vegetables.
Measured in AJDU
Activity of Enzyme: One activity u/mg(u/ml) of pectinase is defined as that quantity of enzyme that can liberate 1 galacturonic acid in one minute at pH of 3.5,
Standard pectinase :1200000 u/ml min
Optimum pH of 3.2~5.0,
The activity of enzymes retains more than 95% when stored at the temperature of 25 Deg C; upto 3 months.
Xylanase
Xylanase (EC 3.2.1.8) is the name given to a class of enzymes which degrade the linear polysaccharide beta-1,4-xylan into xylose, thus breaking down hemicellulose, which is a major component of the cell wall of plants.
As such, it plays a major role in the digestive system of herbivorous micro-organisms (mammals, conversely, do not produce xylanase).
Additionally, xylanases are present in fungi for the degradation of plant matter into usable nutrients.
Commercial applications for xylanase include the chlorine-free bleaching of wood pulp in the papermaking process, and the increased digestibility of silage (in this aspect, it is also used for fermentative composting).
In the future, xylanase may be used for the production of biofuel from unusable plant material.
DIRECT FED MICROBES
Many studies have reported the effective usefulness of DFM in
Detoxifying
Enhancing the immunity System,
Improving F C R
Reducing the Diarrhoea,
The epithelial cells of the intestine are covered by a protective layer of mucus, which is a complex mixture of glycoproteins and glycolipids with the large glycoprotein mucin being the main component. Ability of probiotic bacteria to adhere to the intestinal mucus is considered important for transient colonization, antagonism against pathogens, modulation of the immune system, and enhanced healing of damaged gastric mucosa
Actinomyces, Butyrivibrio fibrisolvens, Fibrobacter succinogenes, Prevotella ruminicola, Ruminococcus albus and Ruminococcus flavefaciens secrete fiber degrading enzymes.
Piromyces rhizinflata secrets carboxymethyl cellulase enzymes.
Bacillus subtilis, Enterococcus diacetylactis, Lactobacillus acidophilus and other Lactobacillus species like L. casei
Would decrease or prevent intestinal establishment of pathogenic microorganisms (Vandevoorde et al., 1991)
Would re-colonize a “stressed” intestinal environment and return gut function to normal more quickly
have reduced incidence of diarrhea (Beecham et al., 1977)
reduced counts of intestinal coliform bacteria (Bruce et al., 1979).
Propionibacteria may be beneficial if inoculated into the rumen (Kung et al., 1991) because higher concentrations of ruminal propionate would be absorbed into the blood and converted to glucose by the liver of the host animal
Acetobacter Xylinum
Degrades levulinic acid.
Production of Dienes, 7 – Cyano Steroids.
Useful in steroid conversion.
Useful in dehydroxylation of cholic acids.
Produces 5 nucleotides by cell culture on hydrocarbons.
Produces 3 keto 1,4 Steroids.
Aspergillus niger
Produces Sachharifying enzymes, glucoamylase, large amounts of maltase, and less amounts of amylase.
Produces Beta galactesidase.
Aspergillus Oryzae
Produces Amylase, Protease, Tannase.
Does not produce aflatoxins.
when fed to lactating ruminants, it is found to result in an average increase in milk production of about 0.45 kg (1.01 lb.) of milk per day.
B lechiniformis
B. polymixa
• Possesses unusual characteristic to fix nitrogen under anaerobic conditions.
• Solubilises Phosphorous.
• Produces polymixin a polypeptide antibiotic, which possess the ability to damage cell membrane structure.
• Asymbiotic Nitrogen fixer.
• Gram negative.
• Produces 2,3 Butanediol used as solvent, humectant, chemical intermediate
B. megaterium
B subtilis
The nutritive qualities of microbial communities that abound the water medium as also possible enhancement of nutritive values of artificial feeds through microbial processing when assessed gave the following results. The crude protein levels (%ges. on dry matter Basis) Bacillus subtilis, Aspergillus flavus, Plankton, Wheat Bran – GNC Mixture, Fermented Bran-Cake , processed Water Hyacinth were 43.63, 47.27. 42.91, 29.09, 36.37 and 14.5respectively. Corresponding Energy values (Kcal / gm dry weight) are 2.98, 4.03, 3.78,3.02, 3.68 and 3.01 respectively. Specific growth rates of the carp fry fed on the above ranged between 3.40 to 0.02. This indicates the significance of microbial communities as triophic components and showed possibilities of their better utilization for carp rearing through medium enrichment and diet incorporation.
• Gram positive. Thermoduric.
• Solubilises Phosphorous.
• Degrades proteins.(Proteolytic) and Carbon (amylolytic).
• Produces amylases and protease enzymes. Used to modify starches, in sizing paper and textiles.
• Can remain active in excreta resulting in less odor, faster decomposition, and in reduction of solids.
• Produces bacitracin, which interferes with regeneration of the monophosphate form of bactoprenol from the pyrophosphate form.
Lactic Acid Bacillus
( Formerly known as Lacto Bacillus Sporogenes )
• Proved effective in lowering LDL Cholesterol.
• Provides an excellent preventative effect against various diseases of the
intestine.
• Increases production of Rotefiers.
• Limits the proliferation of pathogens in rotifiers.
• Provides a source of immunostimulant .
• Useful in the cases of non specific vaginitis / leucorrhoea
• Safe during lactation and in elderly.
Lactobacillus delbrueckii
• Immune Stimulation
• Produces Lactic Acid
L lactis
• Reduces the ability of pathogenic bacteria to grow and cause infection.
• Especially effective against Listeria monorytogenes, which causes severe food poisoning.
L acidophillus
â—¦ Produces extremely effective natural antibiotic substances that can inhibit 11 known disease causing bacteria.
â—¦ It has also been proven to inhibit yeast infections.
â—¦ Helps in cases of chronic constipation and diarrhea by replacing undesirable intestinal organisms.
â—¦ Helps in the cases of food poisoning.
â—¦ Lowers levels of LDL Cholesterol.
â—¦ Found to alleviate intestinal disorders, principle being that the ingestion of large numbers of the lactobacilli may result in replacement of undesirable intestinal organisms by harmless and beneficial organisms, a concept first proposed by the Russian Bacteriologist Metchinikoff in the early days of bacteriology. The implantation of the lactobacilli seems to depend on ingestion of large number of organisms and on supplying a suitable carbohydrate such as lactose that is not readily absorbed by the body but can be easily used by the organism.
â—¦ Aids in nutrient uptake.
â—¦ Fights Candida overgrowth.
â—¦ Controls effectively E Coli and Staphylococcus aureus.
Lactobacillus reutri
• Possess resistance to bile salt and acid.
• Possess the capacity to break down soluble carboxymethyl cellulose, ß-glucan, or xylan. Possess high adhesion efficiency to mucin and mucus.
• Produces an autoaggregation-promoting protein.
• Produces antimicrobial substance reuterin.
• Produces fibrolytic enzymes.
Effective in enhancing the growth and development of poultry subject to stressors similar to those likely to be encountered under commercial production conditions. L.reuri is the only species of known microorganism which can produce reuterin, a newly discovered antimicrobial agent which inhibits growth of other intestinal pathogenic microbes such as Salmonella, Listeria, Escherichia.
In addition, there is evidence that L. reuteri formulations effectively counteract weight losses caused by disease associated stress and competitively reduce Salmonella. In another trial, the result showed that L. reuteri and whey in the diet reduced mortality from natural causes and from mortally induced by a Salmonela challenge.
Also poultry treated with L. reuteri and whey were heavier than non treated animals.
Another conclusion was that L. reuteri and whey in the diets of turkey poults appeared beneficial in the control of caecal Salmonella populations.
Lyophilized L. reuteri exhibit excellent long term viability.
It has been shown that early colonization with L. reuteri contributes to the development of healthy birds, possibly because of the effect it has on the development of the gut mucosal tissues.
Transitory cold stress during the first 48 hours after the placement of the birds causes a growth depression through 20 days of age. Addition of L. reuteri to the diet of these cold stressed birds plays a significant role in overcoming the suppressive effects.
Reduced variability in body weights of the birds was observed with the feeding of L. reuteri resulting in more uniform flock. Controls Cryptosporidium parvum infection. Beneficial in treatment of watery diarrhea.
Sachromyces cerevisiae 3090
• Has ability to improve milk production consistently.
• Helps in oligosaccharide transfer in microsomes.
• May also stimulate rumen fermentation by scavenging excess oxygen from the rumen (Newbold et al., 1996).
• May have a buffering effect in the rumen by mediating the sharp drops in rumen ph, which follows feeding.
• May help to buffer excess lactic acid production when ruminants are fed high concentrate diets.
• Possess invertase activity.
• Produces arginase.
Most investigators agree that yeast culture supplementation strategies can have measurable effects on ruminal fermentations, and a number of beneficial changes in digestion have been noted. Studies in several laboratories have demonstrated that yeast culture supplementation can influence digestive processes in the rumen (Williams and Newbold, 1990; Dawson, 1992; Newbold et al., 1996; Wallace 1996). Typically, the total extent of dry matter digestion was not drastically altered. However, the initial rate of digestion is readily influenced by the addition of live yeast preparations to the diets of ruminants . This is a characteristic of yeast supplementation that has been measured in both in vitro (Dawson and Hopkins, 1991) and in vivo studies (Williams and Newbold, 1990; Smith et al., 1993; Kumar et al., 1997). Since feed intake is often considered to be a function of the initial rates of fiber digestion, early stimulation of ruminal activity can be expected to have a major impact on feed consumption and can provide a driving force for improved animal performance. Such studies suggest an important role for yeast culture supplementation in digestion of animals maintained on high forage diets.
Other studies have demonstrated a role for yeast culture in stabilizing ruminal fermentations and in addressing ruminal disorders. Williams et al. (1991) demonstrated the beneficial effects of the viable yeast culture, on lactic acid concentrations in the rumen in high concentrate diets. In animals fed high energy diets, decreased lactic acid concentrations are associated with higher ruminal pH and are characteristic of much more stable ruminal fermentation. These alterations in ruminal fermentation can be expected to provide for improved digestion and could also be reflected in improved intake and production. The ability of yeast to prevent the accumulation of lactic acid in the rumen suggests a role for viable yeast in overcoming ruminal dysfunctions associated with the use of high energy diets used in both high-producing dairy and fast-growing beef cattle.
Several lines of evidence suggest that yeast culture supplementation can beneficially alter nitrogen metabolism in the rumen . This is reflected in lower ruminal ammonia concentrations observed in animals receiving yeast supplements and is consistent with observed increases in the concentrations of bacteria in the rumen. In addition, these changes are reflected in an increased flow of bacterial nitrogen to the small intestines (Erasmus et al., 1992). Altered nitrogen flow has also been associated with shifts in the basic amino acid flow out of the rumen. The beneficial increase in the flow of microbial protein from the rumen is consistent with models that suggest stimulation of microbial growth in the rumen and more efficient conversion of ammonia nitrogen into microbial protein. Since microbial protein is often used to drive protein synthesis in high-producing ruminants, these observations suggest a role for specific yeast culture supplements in stimulating protein synthesis in both beef and dairy production systems.
Many investigators have attributed the beneficial effects of yeast culture directly to changes in the ruminal fermentation and to changes in the microbial population in the digestive tract . The ability of specific yeast culture preparations to stimulate the growth of ruminal bacteria and to increase the concentrations of specific groups of beneficial bacteria in the rumen has been well documented. Increased concentrations of the total anaerobic bacteria and of cellulolytic bacteria have been one of the most consistently measured responses to yeast culture in the rumen .However, other studies have also suggested that yeast culture preparations can enhance the growth of lactic acid-utilizing bacteria, proteolytic bacteria and bacteria that convert molecular hydrogen to acetate in the rumen. In addition, yeast preparations have been shown to enhance the activities of fiber-digesting fungi in the rumen. Increased concentrations of beneficial microorganisms and enhanced microbial activities can be expected to lead to enhanced digestive processes and the destruction of metabolic intermediates that can result in ruminal dysfunction. The ability of yeast to stimulate specific groups of bacteria is consistent with many of the other physiological and metabolic effects of yeast observed in the rumen and can explain enhanced protein synthesis, improved ruminal stability and improved microbial protein synthesis.
Despite the basic understanding of some of the beneficial effects of yeast cultures on the bacterial population in the rumen, the physiological basis for the enhanced microbial growth has not been completely described. A number of specific hypothetical biochemical mechanisms have been developed to explain the stimulatory effects of yeast cultures in the rumen (Dawson and Girard, 1997). Some of these have been based on the ability of yeast to provide important nutrients or nutritional cofactors that stimulate microbial activities while others suggest that the ability of yeast to control the oxygen level in the ruminal environment is important. These kinds of models have many attractive features but are individually limited in their ability to explain all of the effects associated with yeast supplementation in the rumen.
However, recent studies have suggested that more basic mechanisms are involved in the overall stimulation of beneficial ruminal bacteria. These studies have resulted in the isolation of a group of small compounds that stimulate bacteria to enter into logarithmic growth and thus stimulate microbial activities. Some of the basic characteristics of these stimulatory compounds are consistent with the basic characteristics of small biologically active peptides. The stimulatory activities of these small peptides has been demonstrated in studies with pure cultures of ruminal bacteria. Synthetic tryptophancontaining peptides have also been shown to bring about similar stimulatory effects and have also been shown to stimulate the growth of representative fiber-digesting bacteria from the rumen. These stimulatory activities were not associated with individual amino acids . Stimulatory activities occurred at concentrations well below those that would suggest that these compounds are limiting nutrients. Instead, they appear to serve as metabolic triggers that stimulate beneficial ruminal bacteria to enter into an exponential growth phase. This stimulatory activity toward specific strains of ruminal bacteria can explain many of the observed effects of yeast culture in the rumen.
It is important to recognize that stimulatory effects of viable yeast in the rumen only address some of the beneficial effects of yeast in animal production systems. Research in this area has resulted in a number of new concepts that have evolved into useful animal supplements. Currently yeast cell wall products are available that prevent colonization by pathogenic microorganisms in the gut, alter the composition of microbial populations in the intestinal tract, modulate immune function and alter the structure of the gut wall. These developments are all the result of in depth study of yeast culture supplementation and are currently used to enhance the performance of both ruminant and non-ruminant animals.
Trichoderma reesei
• Produces glucose by enzymatic hydrolysation of cellulose.
• Produces Cellulase.
• Produces D glucanase.
• Produces cell wall lytic enzymes.
When compared to Enzymes, DFM are cost effective and not cumbersome.
However one hurdle in using DFM is that prior to reaching the intestinal tract, these DFM must first survive transit through the stomach, where secretion of gastric acid represents a primary defense mechanism against the majority of ingested microorganisms. BIOOPS has overcome this problem successfully by adopting specific strains that can overcome this problem.
Another problem of using DFM is that nowa days almost all feeds are pelletised at about 105 Deg C, which temperature may damage the DFM. Hence tolerance of DFM to heat is becoming an important factor.
In general, most yeast, Lactobacillus, Bifidobacterium, and Streptococcus are destroyed by heat during pelleting. In contrast, bacilli form stable endospores when conditions for growth are unfavorable and are very resistant to heat, pH, moisture and disinfectants.
BIOOPS has solved this problem by adopting procedures to improve the heat tolerance of the specific strains used.
Organism End Products or Potential Use
Bacillus subtilis amylase, protease
Bifidobacterium bifidum ureases, lactic acid, formic acid
L. lactis amylase, hydrogen peroxide, proteases
Lactobacillus acidophilus lactic acid, acidophilin, glycosidases
Pediococsus acidilactici pediocin (bacteriocin)
Propionibacteria sp. Ruminal lactate utilizer, propionate producer
Propionibacterium thoenii propionicin PLG-1 (bacteriocin)
Bacillus polymyxa Polymixin B Antifungal Peptide
COMPOSITION OF ENZYMIX
Aamylase 325 UI/g
Cellulase 200 Ul/g
Lipase 250 Ul/g
Protease 300 Ul/g
DFM < 1000 Million CFU/g
SALIENT FEATURES OF ZYMEX
• Higher quality end product from cleaner eggs or reduced carcass downgrades
• Improved environment
• improves the digestibility of the feed
• Lower feed costs
• More uniform pigs and birds
• Proven value in antibiotic growth promoter free nutrition
• Provides feed manufacturing with the opportunity to choose low cost feeding stuff to replace high cost feeding stuff.
• Provides feed manufacturing with the opportunity to reduce feed costs
SUGGESTED LEVEL OF INCLUSION:
Generally speaking, adding ZYMEX75- 150 g to 1M T dry materials will play well. But you should reconfirmed suitable dosage depending on your bench-scale experiment results.
OTHER MATTERS OF IMPORTANCE
Use the preparation between pH of 3.8~5.0,
The activity of enzymes retains more than 95% when stored at the temperature of 25 Deg C upto 3 months
WITHDRAWAL PERIOD
Not necessary
STORAGE:
All Enzymes and DFM should be kept away from moisture, excess heat, and light
Ref:
1. Ahmed, F. E. 2003. Genetically modified probiotics in foods. Trends Biotechnol. 21:491-497.
2. Applied and Environmental Microbiology, November 2005, p. 6769-6775, Vol. 71, No. 110099-2240/05/$08.00+0 doi:10.1128/AEM.71.11.6769-6775.2005
3. Baran, M., and V. Kmet. 1987. Effect of pectinase on rumen fermentation in sheep and lambs. Arch. Anim. Nutr. Berlin. 7/8:643.
4. Beauchemin, K. A., W. Z. Yang, and L. M. Rode. 1999. Effects of grain source and enzyme additive on site and extent of nutrient digestion in dairy cows. J. Dairy Sci. 82:378-390.
5. Beauchemin, K., A., and L. M. Rode. 1996. Use of feed enzymes in ruminant nutrition. Proc. of the Canadian Society of Animal Science Annual Meeting, Lethbridge, Alberta. Pp 103-140.
6. Beauchemin, K., A., L. M. Rode, and V.J.H. Sewalt. 1995. Fibrolytic enzymes increase fiber digestibility and growth rate of steers fed dry forages. Can. J. Anim. Sci. 75:641-644.
7. Bedford, M. R., and H. Schulze. 1998. Exogenous enzymes for pigs and poultry. Nutr. Res. Rev. 11:91-114.[CrossRef]
8. Beecham, T. J., J. V. Chambers, and M. D. Cunningham. 1977. Influence of Lactobacillus acidophilus on performance of young dairy calves. J. Dairy Sci. 60(Suppl. 1):74. (Abstract)
9. Beharka, A. A., T. G. Nagaraja, and J. L. Morrill. 1991. Performance and ruminal development of young calves fed diets with Aspergillus oryzae fermentation extracts. J. Dairy Sci. 74:4326-4336.
10. Berkow R, ed. The Merck Manual of Medical Information . Home Ed. Whitehouse Station, NJ: Merck Research Laboratories; 1997.
11. Bruce, B. B., S. E. Gilliland, L. J. Bush, and T. E. Staley. 1979. Influence of feeding cells of Lactobacillus acidophilus on the fecal flora of young dairy calves. Oklahoma Anim. Sci. Res. Rep. 207.
12. Carroccio A, Iacono G, Montalto G, et al. Pancreatic enzyme therapy in childhood celiac disease. A double-blind prospective randomized study. Dig Dis Sci . 1995;40(12):2555-2560.
13. Cheng, K. J., S. S. Lee, H. D. Bae, and J. K. Ha. 1999. Industrial applications of rumen microbes. Asian-Australas. J. Anim. Sci. 12:84-92.
14. Choct, A., and G. Annison. 1992. Anti-nutritive effect of wheat pentosans in broiler-chickens: role of viscosity and gut microflora. Br. J. Poult. Sci. 33:821-834.
15. Dawson, K. A., and D. M. Hopkins. 1991. Differential effects of live yeast on the cellulolytic activities of anaerobic ruminal bacteria. J. Anim. Sci. 69(Suppl. 1):531. (Abstract)
16. Dawson, K. A., K. E. Neuman, and J. A. Boling. 1990. Effects of microbial supplements containing yeast and lactobacilli on roughage-fed ruminal microbial activities. J. Anim. Sci. 68:3392-3398.
17. Ehrmann, M. A., P. Kurzak, J. Bauer, and R. F. Vogel. 2002. Characterization of lactobacilli towards their use as probiotic adjuncts in poultry. J. Appl. Microbiol. 92:966-975.[CrossRef][Medline]
18. Englyst, H. N., S. A. Bingham, S. A. Runswick, E. Collinson, and J. H. Cummings. 1989. Dietary fibre (non-starch polysaccharides) in cereal products. J. Hum. Nutr. Diet. 2:253-271.
19. Feng, P., C. W. Hunt, W. E. Julien, K. Dickinson, and T. Moen. 1992. Effect of enzyme addition on in situ and in vitro degradation of mature cool-season grass forage. J. Anim. Sci. 70(Suppl. 1):309. (Abstract)
20. Fuller, R. 1989. Probiotics in man and animals. J. Appl. Bact. 66: 365-378
21. Gusils, C., O. Oppezzo, R. Pizarro, and S. Gonzalez. 2003. Adhesion of probiotic lactobacilli to chick intestinal mucus. Can. J. Microbiol. 49: 472-478.[CrossRef][Medline]
22. Heck AM; Yanovski JA; Calis KA. Orlistat, a new lipase inhibitor for the management of obesity. Pharmacother . 2000 Mar;20(3):270-279.
23. Hirstov, A., T. A. McAllister, and K. J. Cheng. 1998. Effect of dietary or abomasal supplementation of exogenous polysaccharide-degrading enzymes on rumen fermentation and nutrient digestibility. J. Anim. Sci. 76:3146-3156.
24. Hutchenson, D. P., N. A. Cole, W. Keaton, G. Graham, R. Dunlap, and K. Pitman. 1980. The use of living, nonfreeze-dried Lactobacillus acidophilus culture for receiving feedlot calves. Proc. West. Sec. Amer. Soc. Anim. Sci. 31:213. (Abstract)
25. Jaquette, R. D., R. J. Dennis, J. A. Coalson, D. R. Ware, E. T. Manfredi, and P. L. Read. 1988. Effect of feeding viable Lactobacillus acidophilus (BT1386) on the performance of lactating dairy cows. J. Dairy Sci. 71(Suppl. 1):219. (Abstract)
26. Jenny, B. F., H. J. Vandijk, and J. A. Collins. 1991 Performance and fecal flora of calves fed a Bacillus subtilis concentrate. J. Dairy Sci. 74:1968-1973.
27. Jin, L. Z., Y. W. Ho, N. Abdullah, and S. Jalaludin. 1998. Acid and bile tolerance of Lactobacillus isolated from chicken intestine. Lett. Appl. Microbiol. 27:183-185.[CrossRef][Medline]
28. Kerovuo, J., and S. Tynkkynen. 2000. Expression of Bacillus subtilis phytase in Lactobacillus plantarum 755. Lett. Appl. Microbiol. 30:325-329.
29. Konig, J., R. Grasser, H. Pikor, and K. Vogel. 2002. Determination of xylanase, ß-glucanase, and cellulase activity. Anal. Bioanal. Chem. 374: 80-87.[CrossRef][Medline]
30. Kopency, J., M. Marounek, and K. Holub. 1987. Testing the suitability of the addition of Trichoderma viride cellulases to feed rations for ruminants. Zivocisna Vyroba. 32:587.
31. Kung, L., Jr., and A. O. Hession. 1995. Altering rumen fermentation by microbial inoculation with lactate-utilizing microorganisms. J. Anim. Sci. 73:250-256.
32. Kung, L., Jr., E. M. Kreck, R. S. Tung, A. O. Hession, A. C. Sheperd, M. A. Cohen, H. E. Swain, and J.A.Z. Leedle. 1996. Effects of a live yeast culture and enzymes on in vitro ruminal fermentation and milk production of dairy cows. J. Dairy Sci. 80:2045-2051.
33. Martin, S. A., and D. J. Nisbet. 1992. Effect of direct-fed microbials on rumen microbial fermentation. J. Dairy Sci. 75:1736-1744.
34. Mathlouthi, N., M. A. Mohamed, and M. Larbier. 2003. Effect of enzyme preparation containing xylanase and ß-glucanase on performance of laying hens fed wheat/barley- or maize/soybean meal-based diets. Br. Poult. Sci. 44:60-66.[CrossRef][Medline]
35. Newbold, C. J. 1995a. Microbial feed additives for ruminants. In: Biotechnology in Animal Feeds and Animal Feeding. R. J. Wallace and A. Chesson (Eds.). VCH. New York. Pp. 259-278.
36. Newbold, C. J., R. J. Wallace, and F. M. McIntosh. 1996. Mode of action of the yeast Saccharomyces cerevisiae as a feed additive for ruminants. Brit. J. Nutr. 76:249.
37. Newbold, C. J., R. J. Wallace, X. B. Chen, and F. McIntosh. 1995b. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial numbers in vitro and in sheep. J. Anim. Sci. 73:1811-1818.
38. Orr, C. L., D. R. Ware, E. T. Manfredi, and D. P. Hutchenson. 1988. The effect of continuous feeding of Lactobacillus acidophilus strain BT1386 on gain and feed efficiency of feeder calves. J. Anim. Sci. 66(Suppl. 1): 460. (Abstract)
39. Patterson, J. A., and K. M. Burkholder. 2003. Application of prebiotics and probiotics in poultry production. Poult. Sci. 82:627-631.[Abstract/Free Full Text]
40. Physicians' Desk Reference . 55th ed. Montvale, NJ: Medical Economics Company, Inc.; 2001.
41. Reid, G., and R. Friendship. 2002. Alternatives to antibiotic use; probiotics for the gut. Anim. Biotechnol. 13:97-112.[CrossRef][Medline]
42. Rode, L. M., W. Z. Yanf, and K. A. Beauchemin. 1999. Fibrolytic enzyme supplements for dairy cows in early lactation. J. Dairy Sci. 82:2121-2126.
43. Rojas, M., and P. L. Conway. 2001. A dot blot assay for adhesive components relative to probiotics in microbial growth in biofilms. Methods Enzymol. 336:389-402.[Medline]
44. Roos, S., and H. Jonsson. 2002. A high-molecular-mass cell-surface protein from Lactobacillus reuteri 1063 adheres to mucus components. Microbiology 148:433-442.[Abstract/Free Full Text]
45. Savage, D. C. 1987. Microorganisms associated with epithelial surfaces and the stability of the indigenous gastrointestinal microflora. Die Nahrung. 5-6:383.
46. Scheirlinck, T., J. Mahillion, H. Joos, P. Dhaese, and F. Michiels. 1990. Integration and expression of -amylase and endoglucanase genes in the Lactobacillus plantarum chromosome. Appl. Environ. Microbiol. 55:2130-2137.
47. Selinger, L. B., C. W. Forsberg, and K.-J. Cheng. 1996. The rumen: a unique source of enzymes for enhancing livestock production. Anaerobe 2:263-284.
48. Shils ME, Olson JA, Shike M, eds. Modern Nutrition in Health and Disease . 9th ed. Philadelphia, Pa.: Lea and Febiger; 1999
49. Suarez F, Levitt MD, Adshead J, Barkin JS. Pancreatic supplements reduce symptomatic response of healthy subjects to a high fat meal. Dig Dis Sci . 1999;44(7):1317-1321.
50. Treacher, R. J. and C. W. Hunt. 1996. Recent developments in feed enzymes for ruminants. Proc. Pacific Northwest Nutrition Conference. Seattle, WA.
51. Tricarico, J. M., and K. A. Dawson. 1999. Effects of defined xylanase and cellulase enzyme preparations on digestive processes of ruminal microbial cultures. J. Dairy Sci. 77(Suppl. 1):252. (Abstract)
52. Vandevoorde, L., H. Christianens, and W. Verstraete. 1991. In vitro appraisal of the probiotic value of intestinal lactobacilli. World. J. Microbiol. In addition, Biotechnol. 7:587-592.
53. Varel,V.H., K. K. Kreikemeier, H.J.G. Jung, and R. D. Hatfield. 1993. In vitro stimulation of forage fiber degradation by ruminal microorganisms with Aspergillus oryzae fermentation extract. Appl. Environ. Microbiol. 59:3171-3176.
54. William V Dashek (1997). Methods in Plant Biochemistry and Molecular Biology. CRC Press. ISBN 0-8493-9480-5. p. 313 Google Print reference "Xylans can by hydrolyzed by beta-xylanase"
55. Yang, W. Z., K. A. Beauchemin, and L. M. Rode. 1999. Effects of an enzyme feed additive on extent of digestion and milk production of lactating dairy cows. J. Dairy Sci. 82:391-403.
56. Yanovski SZ, Yanovski JA. Obesity. N Engl J Med . 2002; 346:591-602.